Правка 2008-03-16
Версия 2004-09-16
4. Конкретизация физико-химического уровня мотивационной сферы кратким представлением функций нейромедиаторов
В 1970 годах в исследовании биохимии высшей нервной деятельности произошла революция. В эти годы был открыт ряд процессов управления в коре головного мозга: генерация клетками головного мозга и нейроклетками ряда других органов – специальных веществ, выполняющих функцию сохранения, поддержания и использования информации. Эти вещества обозначаются терминами «нейромедиаторы» (передатчики или neuriotransmitters) и «нейромодуляторы» (усилители или ингибиторы – средства торможения связи, или обобщенно, регуляторы).
Первые сообщения о том, что творческий процесс у человека поддерживается или сопровождается выделением в центральной нервной системе веществ опиоидного характера – эндорфинов и энкефалинов – не мог остаться без внимания в работе, посвященной потребности в творчестве. Этого вполне достаточно, чтобы предположить возможность формирования и существования биологических потребностей (или подкрепления) творчества и проявить интерес к этой области хотя бы на самом общем уровне, какой позволяет высшее даже техническое образование.
Мы не будем вторгаться в сложную сферу биохимии организма,
отметив поверхностно только практическую, или скажем, инженерную, сторону
результатов биохимических исследований. Для читателей технического и
рационального склада, имеющего опыт работы с компьютером и локальными сетями,
значение новации можно «смоделировать» в виде схемы из трех частей.
В первой мы излагаем самое общее представление о нейронной связи.
Во второй мы связываем эти знания с теорией потребностей и мотивации на макроуровне и предлагаем гипотезу метапотребностей с учетом нейромедиаторного механизма межнейронной связи.
В третьей части показываем новое значение и звучание известных прежних утверждений и фактов в свете полученных результатов и предположений.
Четвертая часть объясняет значение ранее казавшихся неясными некоторые природные явления и даже предрассудки, имеющие определенную как выясняется почву.
Предваряя вопрос
читателя – а зачем все это мы читаем, сразу предупрежу, что эксурсом в
нейрофизиологию мы углубляемся в механизм связи между нейронами, в порядке
осуществления которой (связи) и возможно будет увидеть (правда в порядке
гипотезы) то, что нас так волнует в части приобретенных индивидуально
потребностей – 1) запоминание долговременное; 2) реализацию связывания
информации между объектами запоминания, т.н. ассоциативную связь, 3)
осуществление памяти – вспоминание и 4) самое существенное, в некоторых случая
(именно в метапотребностях) самопроизвольное вспоминание того, что нам хотелось
бы повторить.
Центральная нервная система и синаптическая нейромедиаторная связь
Центральная
нервная система (ЦНС) состоит из развитой нервной ткани – сети множества
клеток, именуемых нейронами. Их функция – обработка
информации, включая, вероятно, и хранение памяти в больших нейронных комплексах,
и принятие «решений» через взаимодействие таких комплексов. Количество нейронов
достигает от 5 до 20 млрд. нейронов. Нейроны или, что более вероятно, соединения между ними механизмы
организации соединений являются носителями информации, а сами хранение,
передача и анализ информация и многообразное управления организмом в целом
осуществляется системой, передач почти мгновенных электрических
(электрохимических) взаимодействий между нейронами как между собой, так и между
нейронами, мышцами тела и различными органами ощущений – рецепторами.
Нейроны делят на три группы. Это локальные нейроны, исполняющие функции
общего т. н. «неспецифического» мышления. Это далее сенсорные нейроны, которые связываются с внешними специфическими
(зрение, слух и.п.) системами восприятия через рецепторы (датчики) этих органов.
И это двигательные нейроны, которые
передают управляющие импульсы от центральной нервной системы на органы движения.
Мозг содержит и другие нервные клетки – глиальные, их
на порядок больше нейронов. Глия образует структуру нервной ткани как каркас, и
выполняет, в частности, функции энергоснабжения (метаболические) нейронов.
Нейрон, как и все
другие клетки, окружен полупроницаемой (плазматической, т.е. жидкостной,
биологической) мембраной. От тела нейрона отходят два типа отростков – дендриты (их сотни и тысячи у одного
нейрона) и аксон (один). У большинства нейронов много ветвящихся дендритов,
обеспечивающих входную информацию, но лишь один аксон, выводящий выводной
импульс на другие нейроны. Дендриты обычно коротки, а длина аксона может
составлять от нескольких сантиметров до нескольких метров. Это не удивительно,
если учесть, что у некоторых растений длина молекулы ДНК в хромосоме составляет
до 40 м. Тело нейрона содержит ядро и другие органеллы (биохимические разной
сложности белковые блоки), такие же, как и в других клетках тела.
Морфологическая связь между нейронами. Как же связаны между собою нейроны? Резко выступающая часть нейрона – аксон, его концевой отдел - может ветвиться. Он выходит на дендрит нейрона-соседа или на само тело нейрона-получателя. При этом аксон образует множество окончаний, контактирующих с дендритом следующего нейрона через узкую щель – «синапс» (по-гречески – «соединение»); окончания аксона образуют т. н. «концевую пластинку» или пресинаптическую мембрану – краевую оболочку нейрона-отправителя. У дендрита нейрона-получателя образуется постсинаптическая мембрана. И между пресинаптической мембраной (аксона) нейрона-отправителя и постсинаптической мембраной (дендрита) нейрона-получателя имеется т. н. «межсинаптическая щель».
Синапсы – места связи – могут находиться в разных точках, на телах дендритов или на их выступах, на теле нейрона и на концах аксона. На дендритах и, в меньшем числе, на теле клетки нейрона имеются тысячи синапсов – соединений от внешних нейронов; именно через синапсы аксон, несущий информацию от тела нейрона-отправителя, передает импульс дендритам других нейронов. Синапсы бывают разных принципов действия – электрического (как простейшие по структуре, по большей части для беспозвоночных и низших животных – шириной до 3 нм, т.е. 3*Е10-9 м), химического происхождения (преобладающие для млекопитающих – ширина до 50 нм) и смешанные.
Мы будем рассматривать наиболее важные синапсы химического принципа. Они обеспечивают:
- одностороннее проведение сигнала;
- усиление или торможение (ослабление) сигнала;
- конвергенцию различных сигналов на одной постсинаптической клетке;
- пластичность передачи сигналов (обучение, память и т. д.).
Итак, нейроны – связаны через синусные пространства –
межсинаптические щели. Синусы электрохимическим способом могут обеспечить
одностороннюю проводимость кратковременного электрического сигнала. Таким
образом, связь в межсинаптической щели осуществляется с помощью импульсов электрохимического
происхождения.
Электроимпульсный
характер связи. А теперь обратимся к
вопросу о происхождении импульсов. Общая информация об их зарождении такова. Передача
информации в мозге, как и в нервной системе в целом, осуществляется посредством
нервных импульсов или кратковременных потенциалов «действия» (электрического
происхождения или хемогенных, т.е. электрических импульсов, производимых на
основе высвобождения потенциала в результате химической реакции).
Нервный импульс обычно зарождается в дендритах
нейрона-получателя, специализирующихся на получении информации от других
нейронов, часто сенсорных, и при
передаче импульса телу нейрона.
Химическая основа передачи импульса через синапс. В окончании аксона, которое образует пресинаптическую часть синапса, содержатся маленькие пузырьки (т.н. «везикулы») с нейромедиатором. Нейронная теория, разработанная Рамон-и-Кахалом, знаменитым испанским гистологом, подтверждена биохимически. Ее выводы таковы.
Нейрон, его аксон синтезируют особое вещество нейромедиатор. Он создается в особых пузырьках или везикулах в теле нейрона, которые являются специальными оболочками, не дающими медиатору начать действовать немедленно и не там, где это необходимо. Везикулы заполняются молекулами медиатора и постепенно транспортируются вдоль аксона к нервному окончанию – пресинаптической мембране – месту их будущего применения. Там они накапливаются и хранятся прикрепившись к пресинаптической мембране. В одном окончании аксона могут быть тысячи везикул, каждая размером 40-200 нм. Везикула содержит от 10 тыс. до 100 тыс. молекул медиатора. Под воздействием нервного импульса, приходящего в нервное окончание, эти пузырьки-везикулы, разрываются и изливают своё содержимое в межсинаптическую щель.
О составе
медиаторов и модуляторов в аксоне. Окончание
аксона может содержать несколько типов нейромедиатора, часто в сочетании с
одним или несколькими типами нейромодуляторов. Нейромедиатор, выделившийся из
пресинаптической мембраны аксона, позволяет обеспечить передачу потенциала
действия на рецепторы дендритов постсинаптического нейрона. Мозг использует
разнообразные нейромедиаторы, каждый из которых связывается со своим особым
рецептором.
Таковы выводы теории.
О метаболизме медиаторов. Мы можем еще подробнее остановиться на зарождении и жизненном цикле медиаторов, которые используются при проведении импульса. Наша тема требует внимательного отношения к данным о появлении и функциях (метаболизме) медиаторов, в частности и об их формировании.
Пузырьки (везикулы) – один из стандартных способов хранения и транспортирования активных биологических веществ не только в нейронах, но в любой внутриклеточной среде, с целью сохранить их активность до необходимого момента активации. В нейроне везикулы с медиаторами имеют четыре фазы своей жизни: зарождение (биогенез), транспортирование (например, из центра нейрона к концу аксона) и созревание, выделение из везикул (экзоцитоз) в момент прохождения импульса по аксону и для передачи к нейрону получателю и их быстрое уничтожение (преобразование) или их быстрый сбор, изъятие из межсинаптической щели и восстановление (регенерацию) обратно в везикулы (эндоцитоз). Следует сказать, что для любого инженера, знакомого с электроникой такая функциональность, обеспечивающая импульсный характер передачи и на очень короткое время является просто откровением, впечатляет не только выявленным результатом как творением природы, но и самим фактом и результатом исследования, указывая на сложность труда исследования..
Биогенез нейромедиатора состоит из двух этапов – образования в теле нейрона из специальной генерирующей белки ткани (эндоплазматического ретикулума) пузырьков (везикул), которые затем объединяются специальным внутриклеточным процессом («Гольджи») в «транспортный пакет» - секреторную гранулу-везикулу. На втором этапе ведется передача этих гранул (мембранных образований) по аксону в пресинаптическое окончание.
Созревание везикул-синаптических пузырьков происходит в пресинаптическом окончании и включает два процесса: регенеративное заполнение пузырька местным медиатором (т. н. эндоцитоз или сбор и возврат уже рассеянных в щели молекул медиатора) и прикрепление пузырька к стенкам аксона и к пресинаптической мембране. В заполнении пузырька участвуют, в частности, специфические белки-переносчики медиаторов.
- Экзоцитоз, т.е. рабочее излучение материала из везикул, идет под воздействием электрохимического импульса, катящегося по аксону и под влиянием выделенных ионов кальция (из глии под воздействием импульса). Такое выделение обеспечивается тремя последовательными реакциями:
- образованием контакта между мембраной пузырька и пресинаптической мембраной;
- сцеплением, во время которого белки, участвующие в излучении (экзоцитозе), выстраиваются в определенном порядке и активируются;
- слиянием двух мембран (мембраны везикулы и мембраны стенки аксона или пресинаптической мембраны), т. е. образованием жидкостного электропроводимого контакта.
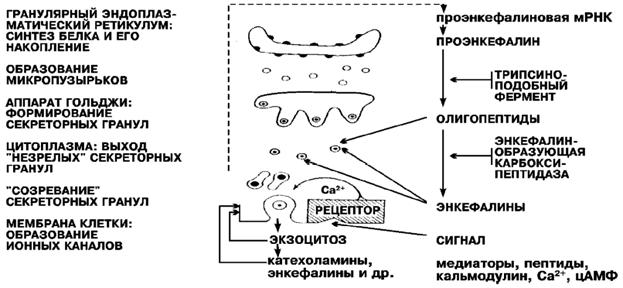
Рис. 4-1(20). Схематическое представление бииогенеза медиаторов [Гомазков О.А., 1999] и его комментарий:
«Последовательные фазы образования секреторных пузырьков — везикул в клетке и этапы биосинтеза энкефалинов. Биохимический (синтез предшественника и его гидролиз) и морфологический (формирование везикул) процессы в секреторной клетке строго согласованы по времени. Биогенез нейропептида регулируется внешними химическими воздействиями (медиаторы, ионы и др.) через рецепторы, локализованные в клеточных мембранах». [Гомазков О.А.,1999]
Теперь о специализации по медиаторам. Каждый
нейрон обычно обладает только таким биохимическим "аппаратом", какой
ему нужен для синтеза конкретных медиаторов, специфически выделяемых из всех
окончаний его аксона. Молекулы медиатора синтезируются путём соединения
предшественников или их изменений в результате ряда ферментативных реакций,
которые управляются из ядра нейрона. Существуют
различные медиаторы, они оказывают на постсинаптический нейрон, т.е. на
нейрон-получатель, либо возбуждающее, либо тормозное (тормозящее) действие.
Передача
отдельного импульса через синаптическую
мембрану. Как и при работе в мембранах
других клеток, передача импульса производится химико-биологическим ионным
механизмом. С чувствительными ионно-химическими датчиками (рецепторами) на
дендритах соединены жидкостные каналы в полупроницаемой постсинаптической
мембране. Эти каналы контролируют движение жидких ионов через мембрану. В покое
нейрон обладает определенным электрическим потенциалом покоя. Как и у всех
мембранных оболочек обычных клеток, разные стороны мембраны (наружная и
внутренняя сторона клетки) имеют разный потенциал. Изнутри клетка нейрона
заряжена отрицательно по отношению к наружной среде. Возбуждение в момент
импульса нейрона-получателя (дендрита – постсинаптической мембраны) реализуется
как усиление потока определенных ионов, главным образом натрия и калия, через
мембрану. В результате отрицательный заряд внутренней поверхности нейрона-получателя
уменьшается – происходит деполяризация – внутренняя стенка мембраны становится
положительной и импульс заряда переходит к (дендриту) нейрону-получателю. Последующее
тормозное влияние (остановка импульса) осуществляется в основном через
изменение потока калия и хлоридов, в результате отрицательный заряд внутренней
поверхности нейрона снова становится больше, чем в покое, и происходит
гиперполяризация. Импульс средствами нейрона-получателя быстро гасится (но
информация снаружи в нейрон-получатель уже поступила).
Еще одним интересным свойством обладает механизм импульсного связывания межсинаптического контакта. Это свойство порционной аннигиляции самого медиатора, обеспечившего контакт. Как только молекула медиатора свяжется со своим рецептором, ее действие (импульс) передается дальше (нейрону-получателю). Параллельно сам носитель – молекула медиатора – уничтожается (инактивируется) другими веществами (аминопептидазами). Биологическая система как бы работает порционно и фиксировано во временном интервале во избежание слишком длительного действия и нарушения точного контроля передачи.
Ионный механизм
возникновения импульса при интеграции потенциалов в нейроне.
А теперь обратимся к интегративной функции нейрона. На
входе нейрона, как мы говорили, тысячи дендритов (входов) и один аксон (выход).
Одна из функций нейрона состоит в интеграции (суммировании) всех воздействий,
воспринимаемых от других нейронов через синапсы на его теле и на дендритах.
Поскольку эти влияния могут быть возбуждающими или тормозными и могут не
совпадать по времени, нейрон должен биологически, точнее, электрохимически,
аккумулировать (с позиций информатики - исчислять) общий эффект синаптической (входной,
внешней) активности (или воздействий всех своих нейронов – излучателей,
соседей) как функцию времени. И результатом такой интеграции в зависимости от
настройки данного нейрона может быть один единственный импульс на выходной
аксоне. Таким образом, по своим функциям каждый нейрон многократно сложнее
простой системы хранения памяти, например, в компьютере и для сравнения
мощности современных компьютеров с нейронной сетью человека до выяснения
механизмов аккумуляции входной информации в нейроне пока нет достаточных
оснований.
Продолжим. В покое нейрон обладает (отрицательным) электрическим
потенциалом покоя (в 70 милливольт) по отношению к наружной мембране. Если
возбуждающее действие внешних импульсов преобладает над тормозным действием, и
деполяризация мембраны превышает пороговую величину, происходит активация
определенной части мембраны нейрона – в области основания его аксона (аксонного
бугорка). Здесь в результате открытия каналов для ионов натрия и калия
возникает потенциал действия (нервный импульс положительного знака). Этот
электрический потенциал распространяется далее по аксону к его окончанию.
Выходной импульс, таким образом, оказывается результатом
фильтрующего влияния данного нейрона при его пропускании через себя сотен и
многих тысяч других импульсов, их «пачек» - последовательностей.
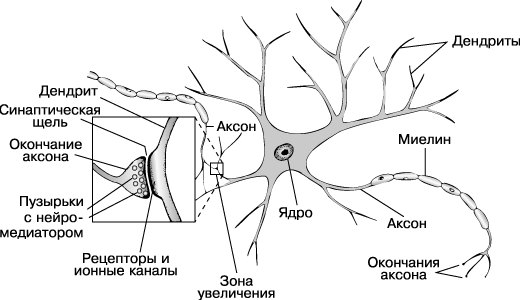
Рис. 4-2(21). Нейрон и соединение его с аксоном нейрона – соседа (слева), воспроизводится по по http://www.krugosvet.ru/articles/38/1003886/0003637G.htm
Движение по аксонам и дендритам. Само движение импульсов по проводящим путям тоже достойно внимания. Многие аксоны покрыты особой (миелиновой) оболочкой, которая
образована многократно закрученной мембраной глиальных клеток. Благодаря этому
скорость проведения импульса (или потенциала действия) по аксону может
увеличиваться на порядок, достигая многих метров в секунду.
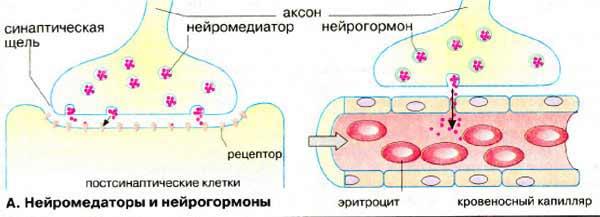
Рис 4-3(21а). Нейрон и соединение его с другими нейронами. Справа изображены более медленно действующие механизмы управления нейронами организма – выделение в кровь нейрогормонов, которые иерархически выше нейромедиаторов и имеют интервалы действия, измеряемые часами и сутками, по [Кольман Я., Рем К.-Г.]
Итак, теперь мы может подвести итог. Медиатор генерируется, передается на место использования, срабатывает от потенциала действия или импульса и передает импульс через синапс и затем аннигилирует (теряет активность) с помощью ряда различных химических процессов, протекающих на рецепторах нейрона - получателя, подготавливая систему к работе по приему нового импульса. Кроме того, разрушенный медиатор в ряде процессов снова восстанавливается и возвращается в везикулы для многократного использования.
Действие медиаторов и дополнительных нейромодуляторов может быть ускоряющим и усиливающим, и наоборот, (и реже) замедляющим и гасящим. Множество различных медиаторов и модуляторов образуют сложные комбинации, в которых могут участвовать те же составляющие, но в разных пропорциях и как результат с противоположным или резко различным эффектом. Кроме того, следует учитывать, что, кроме собственно медиаторов и модуляторов с пресинаптической стороны существует управляемая биохимическая среда на стороне получения сигнала – за постсинаптической мембраной, возле рецепторов нейрона-получателя, воспринимающих потенциал действия.
Нейромедиаторные и модуляторные, а также нейротормозные функции, несомненно, имеют свою иерархию, которая разворачивается во времени и завершается на более медленно функционирующем нейрогормональном уровне.
В настоящее время механизм связывания нейронов через медиаторную транссинаптическую технологию можно считать общепризнанным.
Квантовая теория Бернарда Катца (1953) и везикулярная теория - гипотеза освобождения медиатора в синапсе дополняют друг друга. Мы особенно отмечаем важную для наших будущих предположений деталь этих теорий. В квантовой теории указывается на возможность самопроизвольного излучения порций медиаторов из везикул, как это представлено, например, А. Л. Зефировым (Статьи Соросовского образовательного журнала в текстовом формате), ниже приведен текст Зефирова:
«Бернард
Катц с сотрудниками, исследуя нервно-мышечный синапс лягушки (медиатор -
ацетилхолин), с помощью микроэлектродов зарегистрировали новый класс
постсинаптических потенциалов. Эти сигналы возникали случайно, в покое, имели
очень маленькую амплитуду и были названы миниатюрными потенциалами концевой
пластинки (МПКП). Далее оказалось, что постсинаптические потенциалы, вызванные
раздражением двигательного нерва (потенциалы концевой пластинки - ПКП), от
раздражения к раздражению варьируют по амплитуде, причем эти колебания кратны
амплитуде МПКП. Было предположено, что медиатор в синапсе освобождается в виде
мультимолекулярных порций - квантов. В покое случайное освобождение из
нервного окончания отдельных порций вызывает появление на постсинаптической
мембране МПКП, а в ответ на раздражение происходит синхронное освобождение
нескольких десятков или сот квантов и возникает ПКП.(мое выделение - СЧ).
Электрофизиологическое определение показало, что квант медиатора состоит из
1000 - 10000 молекул ацетилхолина. В дальнейшем квантовая гипотеза освобождения
медиатора получила подтверждение на самых разнообразных объектах и в настоящее
время считается общепризнанной.»
[Зефиров А.Л. , 2000]
Здесь авторов волнует дискретность и порционность сигналов, нас же
в этом интересует сам факт случайного освобождения медиатора в синапсе «в
покое».
Теперь, завершив краткий анализ, мы можем начать восходящее
движение к синтезу нашей гипотезы.
Попытка синтеза. Нервные импульсы
поступают от периферических органов тела на сенсорные нейроны. Локальные
нейроны получают такие импульсы от сенсорных и двигательных нейронов и
собственно от других локальных нейронов, наконец, двигательным нейронам
«приказы» поступают от локальных нейронов. Кроме того, поток импульсов ведется
одновременно от различных нейроисточников, и для конкретных нейронов, основная часть потока
импульсов является шумовым. Кроме того, как отмечено выше, существует или
возможно шумовое выделение отдельных порций медиаторов возле пресинаптической
мембраны, что как теперь подтверждено (Б. Катц, А. Л. Зефиров), может создавать
отдельные самогенерации импульсов и пачек импульсов.
Электрические импульсы поступают с различными частотами, часто
пачками или сериями как процессы возбуждений и могут передаваться с
определенной частотой через другие нейроны. Обычно нейроны «работают», т.е.
возбуждаются, целыми ансамблями. Установлено, что в некоторых состояниях передача
сигналов от одной группы нейронов к другой осуществляется на определенной
частоте, которая служит информативным параметром для распознавания.
Нейрон-получатель реагирует не на весь многочастотный поток поступающих
импульсов извне, а на пачку импульсов вполне определенной частоты, которая и является в его, нейрона,
«понимании» сигналом, несущим информацию.
На входе нейрона сотни и тысячи
электрических сигналов от других нейронов, в том числе и сигналов,
представляющих частично и шумовые импульсы (данные). Выходной сигнал на аксоне
выступает как интегральный результат многих сот внешних входных импульсов. Это
электрический импульс (в реальности это пачки импульсов различной частоты),
который является логическим решающим или выходным сигналом (по П. К. Анохину)
нейрона на внешнее множественное возбуждение [Анохин П. К., 1974, с. 4, Анохин
П. К., 1979, с. 54-55]. Это означает, что нейроны образуют собою гигантскую информационную сеть,
структурированную своими настройками из медиаторов и модуляторов, как фильтры к
входной информации и как частично автогенераторы собственных импульсов –
саморазрыв везикул. В этой сети по мнению многих исследователей нейроны
образуют связанные между собою ансамбли или констелляции (дословно – созвездия).
Конфигурация и состав их существенно изменяется при активизации и при различных
формах нервной деятельности. Мы можем предполагать, что образное хранения
информации, связано с такими электрохимическими и нейробиологическими
конструктами (образованиями).
Долговременная память, эмоции и опиоидные нейромедиаторы
Мы можем опираться на функциональное определение памяти, данное академиком И.П. Ашмариным:
«Биологическая память – способность живых существ или их популяций, воспринимая воздействия извне, закреплять сохранять и в последующем воспроизводить вызываемое этими воздействиями изменения функционального состояния и структуры» [Ашмарин И.П., 1987, с. 3].
Это определение можно толковать и как способность объекта – хранителя памяти моделировать (сохранять и позже воспроизводить) состояния, структуру (или динамические процессы их изменения) внешней среды, отраженной определенным образом в объекте.
Полученные самые общие данные о нижнем биохимическом «этаже» памяти позволяют представлять ее как совокупность конкретной конфигурации размещения и постоянно поддерживаемой выработки, воспроизведения нейромедиаторов и модуляторов в ансамблях нейронов. О том, что долговременная память связана с действием опиоидов, указывают результаты исследований 80-х гг., см. например, [Стайн Л., Беллуци Дж., с. 295-298].
Каков потенциальный объем такой памяти? Если предполагать единицей информации связь между нейронами - импульс и его отсутствие, то считать следует до уровня синапса. Если оценивать количество нейронов в размере 20-100 млрд. единиц и предполагать, что синапсы на входе нейронов на три порядка выше, и считать, что синапсы в нейроне-излучателе или нейроне-получателе являются не только элементами связи, но и элементами двоичной памяти, то объем памяти мозга достигает до 20-100 Гбит или 2-12 Гбайт. Но это упрощенно и формально. В реальности вероятно, значительно меньше. Уже неоднократно говорилось о том, что человек не использует все свойства своей памяти. Да, в целом, у нас не имеется базы для сравнения мозга человека с компьютером. Его свойства много ценнее свойств любого компьютера, потому что люди вряд ли будут проектировать компьютер с собственными потребностями и встроенной программой формирования целей борьбы за выживание в окружающей среде (для удовлетворения потребностей и за собственное воспроизведение).
Хорошо сказал Б.Ф. Поршнев о памяти и о фильтрации внешних возбуждений среды в центральной нервной системе
В
сущности, нервное возбуждение - это тоже простая реактивность, хотя на очень
специальном и сложном химико-физическом субстрате. Биологическая эволюция
"отрицает" его (как свободное передвижение животного
"отрицает" силу тяжести), ибо возбуждение все жестче перекрывается
шлюзом, канализируется. "Ум" животного - это возможность не
реагировать в 999 случаях из 1000 возникновении возбуждения. Животное все
успешнее, где только возможно, оберегает себя от реакции, так как движется,
обладает дистантной рецепцией, дробит раздражители с помощью мозговых
анализаторов. Все это не имеет никакого отношения к экономии энергии
реактивности: напротив, расход энергии на торможение растет в ходе эволюции в
гигантских пропорциях. [Поршнев
Б.Ф., 1974, Гл.5].
Теперь, мы прошли путь вниз почти до биохимии и до нанометров и поднялись снова наверх к цели нашего исследования, к системе потребностей, к надстроенным потребностям или метапотребностям. Мы будем говорить о такой памяти на действия, на манеру действия, которая может стать источником удовольствия, наслаждением и самостоятельной целью, и которую мы, поэтому, собираемся именовать метапотребностью. При этом мы рассматриваем уровень не кратковременной памяти в ЦНС, а именно долговременной, которая только и может закрепить, удерживать установки в поведении индивида.
Роль эмоций, вероятно, недооценивают. Так, у Д.В. Колесова – это «гедонистическая» компонента – оценка эффективности удовлетворения потребности. В оценке эмоций видят нечто нефункциональное – лишний довесок к психологии поведения - эмоции накапливаются в памяти, и у человека появляется возможность испытывать эмоции в связи с событиями в прошлом и в «обдумывании чего-либо на будущее», в лучшем случае – эмоция – это «физиологически-мобилизующая» составляющая поведения [Колесов Д.В., 2002, с.122-123]. У нас создается впечатление, что эмоции имеют много более важное регуляторное значение. О роли эмоций, их значении, возможно, лучше всего сказал П.К. Анохин: «Эмоции бы отмерли в отсутствии их функционального значении», а критикуя книгу Э. Гельгорна и Дж. Луфборроу «Эмоции и эмоциональные расстройства», он говорил о «саморегуляторном характере эмоциональных разрядов и о приспособительной роли эмоций» [Анохин П.К., 1979, с. 447-449].
Присоединим к изложенной выше информации все, что мы знаем о роли положительных и отрицательных эмоций и их связи с памятью.
Эмоция как положительное подкрепление и дополнение к удовлетворенной потребности устанавливает и закрепляет память (самое малое, резко улучшает установление и закрепление памяти) об инструментальном поведении, удовлетворяющем потребность. Положительное подкрепление может быть и нейтральным, но при депривациях положительным подкреплением оказывается устранение отрицательного состояния возбуждения, страдания, беспокойства и т.п. При повторении действия, которое является полезным для индивида (или биологической особи), это действие запоминается в ЦНС. При сильной эмоции (пиковом переживании) запоминается сама эмоция и связанное с ней действие.
При отрицательном подкреплении – при не удовлетворении потребности – возникает отрицательная эмоция, в лучшем случае нейтральное состояние в ответ на зря потраченные усилия. В результате (который на уровне психики является эмоцией) данное инструментальное действие или забывается как бесполезное или формируется как опасное негативное и запрещенное к применению или излишне затратное. И его избегают.
Примечание: Из этого следует
важное правило – если вы хотите продолжать усилия при появлении трудностей –
старайтесь не расстраиваться – это условие для мобилизации сил на продолжение
дальнейших попыток. Понятное житейское правило получает теперь строгое научное
основание.
Для нас важно то, что сказал Петр Кузьмич Анохин об эмоциях в части обычных потребностей (голода):
«в структуру акцептора результата действия включается эмоциональный компонент, предвидение той положительной эмоции (курсив мой – СЧ), которая обычно всегда сопровождает удовлетворение доминирующих потребностей. Именно эти процессы эмоционального «предвкушения» составляют основу того общераспространенного состояния, которое в обыденной жизни получило название «аппетит»…», [Анохин П.К., 1979, с.10].
По сути, Анохин сказал более точно о пусковом характере эмоции и даже об эмоции как эмоциональной психологической надстройке над биологической потребностью голода. Но «аппетит» - это только одна индивидуальная потребность или метапотребность гурмана (не чрезмерно сытого любителя тонких вкусовых ощущений) и т.д. и т.п. Мы ищем и, похоже, обнаруживаем ключ к метапотребностям над физиологическими потребностями - предвкушение как объяснение природы пускового процесса.
В изложении проблемы мы столкнулись со следующим не проясненным моментом нашего общего видения. Представим себе, что эмоций нет, что человек как бездушное техническое устройство, удовлетворяет свои технические (ну, скажем, физиологические) потребности (как и заправка топливом или техническое обслуживание). Если мы исключим эмоции как внешнее (психологическое) проявление оценки удовлетворения потребности нашим субъектом (и объектом исследования), то нам необходимо вычленить и найти вместо эмоций новый физиологический аппарат сравнения и оценки результата действия – степени удовлетворения потребности и качества результата в мышлении и в поведении человека. Это означает, что мы пытаемся идентифицировать психологический и физиологический блок сравнения результата и поставленной цели в психике человека, т.е. идентифицируем то, что именуется у Анохина блоком оценочной афферентации.
ПРИМЕЧАНИЕ: Напомним, что вся система Анохина представляет собой символические структуры нашего познания – они, структуры, по идее «системы управления» (в модели кибернетики) должны присутствовать в организме и в мышлении, но мы сами должны найти эти реальные механизмы и подставить их на место схемы Анохина в поле нейрофизиологии.
Нам, людям рациональным, желательно представлять сравнение цели и результата как рациональный блок мышления, сродни решающим блокам в абстрактной системе автоматического управления. На очень высоком уровне сознания мы готовы признать, что человек разумный именно так себя и ведет, в духе теории Альберта Бандуры, – логично сравнивает и логично принимает решение - исключить метод или включить метод в систему своих инструментальных средств. Но у древнего человека, такого аппарата априори не было. В реальности и на подсознании и у зверя с его рефлексами, и у преантропа и у современного младенца есть только эмоция как отражение его оценки физиологического состояния (своеобразной оценки уровня до логики). Она и лежит ниже рационального, и она предваряет работу рационального, более позднего аппарата в сознании. И вообще в жизни каждого человека онтогенетически начало мышления и построения символической системы опирается на эмоции как блок оценочной афферентации. Другого не дано! Потому мы вплоть до серьезного обоснования других содержательных оценок другими авторами в порядке гипотезы рассматриваем следующее: эмоция есть блок (инструмент) низшего уровня психологии в процессе оценочной афферентации.
Имеющиеся данные, например, [Эндорфины, сб.ст. 1981], указывают на выработку в ЦНС в момент активности в животном мире при положительном или отрицательном результате (оценочная афферентация) соответствующих биохимических веществ класса нейромедиаторов и нейромодуляторов.
Поэтому у нас имеется основания предположить логическую связь между фактом возникновения эмоции и перестройки на биохимическом уровне памяти ЦНС как настройки нейронной сети с конкретным механизмом модификации (перенастройки) объема генерации нейромедиаторов и модуляторов. Для удаления условных рефлексов, которые не подтвердили потребность и ожидание ее удовлетворения, начинают «работать» отрицательные эмоции, при этом они формируют «тормозные пути», разрушают или стирают, перенастраивают синапсы конкретного старого медиаторного механизма рефлекса или даже, возможно, более того, создают отвращение к какому-либо действию (определяем его как отрицательно подкрепленное, как фобию). Последнее мы можем рассматривать как новый уже отрицательный условный рефлекс (депривационного типа) – не использовать то-то, не делать то-то, системный запрет – это тоже установка.
А при положительных эмоциях (положительном подкреплении) в ЦНС осуществленная активность фиксируется, или на макроуровне «запоминается», экзоцитозом (выделением) дополнительных порций нейромедиаторов дофаминового типа в системе тех нейронных ансамблей и их взаимодействий, которые функционировали при подкрепленном акте. При этом закрепляется, реформируется и устойчивая биохимическая связь между нейронами. И это уже видел Анохин, утверждая, что:
«Все виды потребностей приобрели побудительный характер,… формирующий различные типы побудительного или … отвергающего поведения» [Анохин П.К., 1979, с. 449].
Итак, на множестве научных оценок и обобщающих мнений мы установили, что
- Результат (успешный или отрицательный) деятельности или активности и(или) сама активность могут воспроизводить соответственно сильные положительные и отрицательные эмоции, т.е. состояния возбуждения
- Эмоциональные состояния способствуют (положительные) запоминанию или (отрицательные) стиранию памяти, т.е. (появления запрета) действий и ситуаций.
- Исключительно все эмоциональные состояния сопровождаются выделением биохимических веществ, часть из которых носит опиоидный характер (часть положительных эмоций).
- Отдельно доказано, что опиаты служат закреплению различных условных рефлексов.
Указанные положения можно представить схемой, из которой следует, что положительные эмоции и выделение некоторых биогенных опиоидов суть одно явление на двух уровнях, см. рис. 4-4(22).
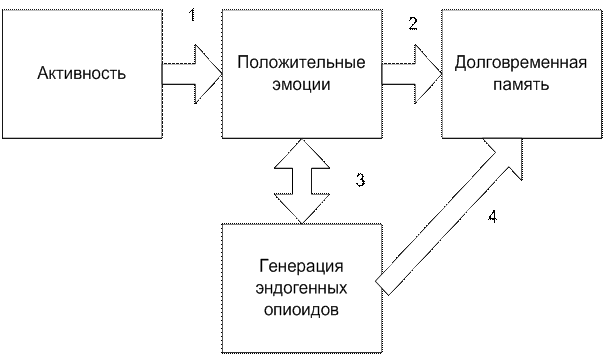
Рис. 4-4(22). Формирование долговременной памяти о полученном положительном подкреплении (условный рефлекс), номера стрелок указывают на установленные в науке связи объектов.
Поскольку мы ищем механизм появления, формирования и проявления (реализации) вторичных или метапотребностей, то нас интересует далее механизм возникновения памяти о прошедших положительных подкреплениях или представление о будущем положительном подкреплении (предвосхищение) как источник последующих действий. Первоначально кажется возможным построить симметричную схему, см. рис. 4-5(23).
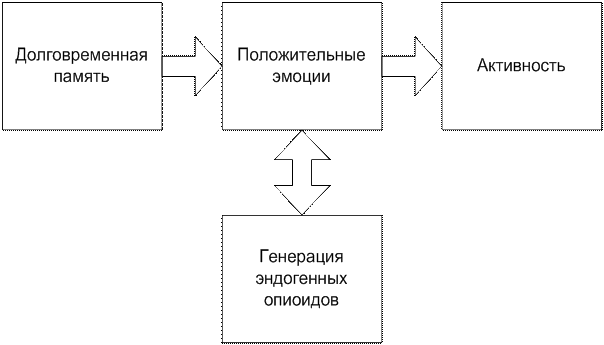
Рис. 4-5(23). Формирование новой активности в связи с воспоминанием о полученном положительном подкреплении (условный рефлекс). Вариант 1.
В этой схеме, рис. 4-5(23)., построенные нейронные связи вместе с эмоциями начинают симметрично работать на активность, запускать действие. Однако, здесь мы не имеем нижнего уровня запуска долговременной памяти – нейронного механизма работы.
Мы останавливаемся и возвращаемся к более простой схеме – схеме обычной потребности, но с нейронным уровнем. Мы представляем процесс формирования установки или условного рефлекса как первичное выделение нейромедиаторов в момент удовлетворения потребности. В результате изменяется настройка нейронной сети. В такой момент и после нескольких повторов происходит существенная реконструкция нейронных констелляций. Аналогично настраиваются условные рефлексы у животных. Последующую реализацию потребности мы строим по достаточно четкому представлению П.К. Анохина и в развитие этого представления, выделив только важные для нас компоненты. Формирование цели и мотива мы заменяем включением структуры «установки», которая уже сформирована и устойчиво служит удовлетворению потребности, см. рис. 4-6(24).
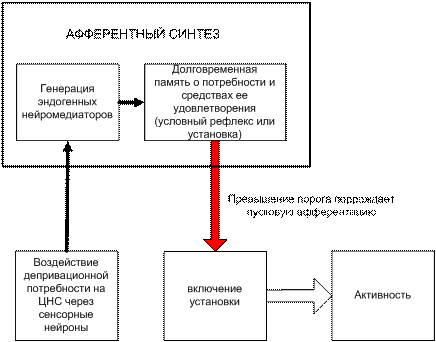
Рис. 4-6(24). Процесс формирования установки под воздействием физиологической потребности (из модели Анохина с учетом теории излучения нейромедиаторных веществ).
При последующем многократном повторении воздействий от сенсорных нейронов, отражающих обычные физиологические депривационные потребности, рис. 4-6(24)., нейроны «настроенных» ансамблей обеспечивают в каждой ситуации выделение нужной комбинации нейромедиаторов и как результат - формирование специфического возбуждения. Последнее отражает и раскрывает в эффекторной системе импульсов уже выработанную установку поведения животного или человека как (мотивированную инструментальную) реакцию на потребность или внешнее раздражение. Итак, нейронный механизм запуска – важнейшее звено, без использования (функции) которого не объяснить начало любого процесса более высокого уровня, уровня психологии.
Несколько отвлечемся и расширим взгляд. Формирование любого простейшего устойчивого инструментального действия при появлении и удовлетворении определенной потребности мы считаем составной частью развития культуры социума. С учетом сказанного мы можем теперь объяснять рост и развитие культуры не только рациональным поведением человека разумного, но, в первую очередь, на основе положительной эмоции, закрепляющей его успехи, и отрицательной эмоции, отстраняющей его от повторения неверных действий и ошибок. Эмоции обеспечивают, таким образом, связь наших низменных и простейших потребностей с высокой технологической культурой. Это и есть ответ и поддержка мнения Анохина о роли эмоций в развитии человека, да и в адаптации биологического развитого мира в меняющемся окружении.
Гипотеза о нейромедиаторном механизме активации метапотребности
Мы возвращаемся к основному направлению нашего анализа: оценить появление потребностей высшего плана, которые не имеют и не могут иметь прямого источника возбуждения от периферической системы. При отсутствии внешнего возбуждения от вегетативной нервной системы ситуация значительно усложняется. У нас нет теперь иного гипотетического источника, кроме текущего внешнего неспецифического окружения и внутреннего источника в центральной нервной системе.
(Примечание:
неспецифичность здесь понимается как отсутствие прямого основания для поведения
– например, – это не угроза от среды, которая вызывает тревогу и т.д.)
Источник раздражения (возбуждения) теперь находится внутри ЦНС. Мы рассматриваем здесь только потребности, возникающие под воздействием сильного эмоционального положительного возбуждения (метапотребности). Слабым аналогом такой метапотребности служит интенсивно изучаемый механизм поддержания наркомании. Однако, мы исключим все противоположные понимания в части обстоятельств появления метапотребностей.
Метапотребность инициируется внутри ЦНС индивида, а не вне его. Рассмотрим роль влияния внешних факторов в инициации метапотребности или ассоциаций памяти (метапотребности) с внешними раздражениями. Внешние события в жизни и человека, и особи в биологическом мире могут ассоциироваться с потребностью и, в том числе, с метапотребностью. Но в целом внешние воздействия подразумевают настроенность индивида или особи на конкретную деятельность или на конкретную потребность (еще раз, мы не обсуждаем различные, явные опасности для особи, индивида – метапотребность – это именно та тема – высшая потребность, которая предполагает озабоченности низкого порядка).
В этой связи можно считать, что интересующее индивида действие будет ассоциировано или вызвано буквально любыми событиями внешней среды. Это хорошо иллюстрируется на бытовом уровне даже для обычных потребностей такими шутками, вроде фразы: «кто о чем, а вшивый о бане». Или это отражается простой солдатской шуткой о том, что солдат всегда думает о женщине: «а я завсегда о них думаю!». Если говорить о голоде, то влияние внешних событий носит буквально символический характер – если потребность существует, то внешние факторы являются, скорее, предлогом и поводом, чем причиной инициации потребности. По сути, большинство внешних раздражителей или воспоминаний начинает ассоциироваться с доминантой в сознании человека. Отсюда же возникают и патологии в части т. н. «навязчивых идей».
И тем самым мы подходим постепенно к выводу о самогенерации потребности. У большинства молодых мужчин случается во сне оргазм и эякуляция при долгом воздержании, но, как правило, не в случае регулярной половой жизни. Поэтому и метапотребность, как и обычные индивидуальные физиологические потребности сильнее ощущается, если она не удовлетворялась некоторое время. Короче, она возникает сама собой – это уже и есть потребность. Если же она удовлетворяется ежедневно (например, игра, творчество и т.п.), то индивид не ощущает ее как потребность – он просто ежедневно занимается тем, что является его удовольствием. Здесь достаточно привести слова известного русского тенора Дмитрия Хворостовского о том, что «Петь – это колоссальное удовольствие» (май 2004).
Таким образом, мы можем определить и орган, потребность которого мы испытываем, или физиологическую функцию органа, которая требует своего удовлетворения в виде метапотребности через запись ее в ЦНС. Но сюда же относятся и неспецифические установки и привычки, которые записаны в ЦНС и подкреплены позитивным ожиданием и поисковой активностью. И потому, то в настоящий момент можно утверждать, что метапотребность не что иное как приобретенная физиологическая потребность или это приобретенная потребность собственно психики, самого сознания индивида (на уровне психологии), записанная в центральную нервную систему (на физиологическом уровне).
Итак, при наличии потребности как внутреннего побуждения внешние обстоятельства не должны играть ведущую роль. Внешние обстоятельства в целом для животного и для человека, для их потребностной сферы, имеют ограниченный диапазон Если господствуют внешние обстоятельства, то мы имеем (по Маслоу) узкую группу потребностей: потребности безопасности 1 (самой жизни) и 2 (надежного обеспечения физиологических потребностей) и социализующую человека потребность в любви и принадлежности. Мы исключим далее различные угрозы при обсуждении темы метапотребности.
Теперь мы полностью концентрируем свое внимание на автовозбуждении. В соответствии с рис. 4-6 нас интересует работа долговременной памяти, а именно, ее автогенерация. Сопоставление объектов низкого - биохимического – уровня (везикулярные эндогенные концентрации) и высокого – психологического - уровня долговременной памяти с высокоуровневой системной моделью афферентации Анохина подводит нас к более точному пониманию порядка возбуждения. Дело в том, что модель реализации или воспроизведения долговременной памяти – это макромодель – в ее основе так же лежат нейрохимические процессы. Как мы знаем из общесистемных представлений в основе формирования макропроцессов высокого уровня всегда лежат микропроцессы, накапливающие возбуждение (или любой другой кумулятивный эффект как переход количества в качество) на более низких структурных уровнях. Поэтому, когда мы говорим о стремлении к гедонистическому поведению (Колесов Д.В.), то нам следует начинать оценивать психологический механизм с действий биохимического уровня. И только потом мы можем говорить об эмоциях, о памяти, о предвкушении положительных ощущений на верхнем уровне.
Когда мы возвращаемся к множеству наблюдений и обобщений специалистов о накоплении нейромедиаторов в везикулах, о генерации импульсов в нейронной сети, мы обращаем внимание на встречающиеся мнения исследователей о самогенерации импульсов, о наличии даже некоего нормального фона и «шума» нейроимпульсов. Этот поток представляет собою множество конкурирующих информационных воздействий от внешней среды и от автогенерации различных нейронных ансамблей. Большая часть таких воздействий не воспринимается сознанием как содержательная информация и отфильтровывается как шумовая тормозными процессами нейросистемы индивида. Нейробиологов и психологов в данной теме обычно больше интересуют процессы торможения и возбуждения. Наше внимание обращено именно на шумовой фон. Из автогенерации мы получаем представление о том, что идущее накопление материала в везикулах может прерываться самопроизвольным разрывом части везикул и спонтанным впрыском части медиаторного материала в межсинаптическое пространство, что и может приводить как к качественной передаче импульса, так и к снижению порога генерации импульса в определенной специфической ситуации.
Именно с этой достаточно общей и устойчивой позиции мы можем формировать представление, что связь между какими-либо явлениями начинает отфильтровываться ансамблями нейронов, настроенных на определенное положительное подкрепление (метапотребность) в пользу данной надстроенной потребности (независимо от состава внешних событий). Но мы отмечаем, что состояние этих ансамблей должны быть «желающими». Нейронные блоки, ансамбли, констелляции должны иметь избыток специфических нейромедиаторов на этот момент.
Отчего и по какой причине мы можем предполагать избыток? Такой избыток складывается от непрерывного процесса удовлетворения, точнее, генерирования, данной потребности индивида в прошлом, или от очень сильного однократного положительного подкрепления потребности (установки, просто действия) в прошлом. Мы имеем в виду одно действие (очень редко) или уже существующую индивидуальную практику и установку. Если организм растрачивал определенное количество нейромедиаторов в процессе жизнедеятельности, то нейронный клетки далее формируют обычный материал, начинают формировать новый материал для будущей работы. Если ожидаемой работы не возникло, материал накапливается в избыточном количестве, пребывая в секреторных контейнерах – везикулах. И тогда начинается спонтанный экзоцитоз медиаторов. Это и есть потребность. Идет фильтрация всей другой информации. Ассоциации возникают только в одном направлении и т. п. Нервная система требует обычного режима их расходования, организм требует удовлетворения важной для него потребности. Схема не сложнее, чем выделение желудочного сока в обычный час обеда. Только потребность может быть настолько мета-, что о ней знает лишь сам ее носитель (например, маньяк).
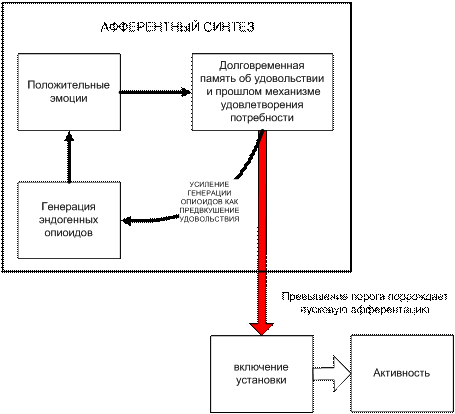
Рис. 4-7(25). Возникновение пусковой афферентации в процессе афферентного синтеза
Положительные метапотребности
Воспоминание при отсутствии деятельности. Итак, если ансамбли нейронов, зафиксировавшие память о положительных результатах или процессах в какой-либо активности, вызрели или накоплены и «готовы» к впрыску, то пороговый уровень внешних импульсных воздействий на ансамбль вызовет автогенерацию собственных возбуждений и спонтанный цепной процесс возбуждения при произвольных внешних воздействиях шумового типа. На уровне психики это можно именовать воспоминанием или спонтанной ассоциацией желания с любым из внешних факторов. Воспоминание или ассоциация с прошлым удовольствием возвращает индивида к представлению об удовольствию и поток извержения нейроопиоидных пептидов в мозге может нарастать по представлению – это и есть «предвкушение» («аппетит» и т.п.). На рис. 4-7. это изображено петлей обратной связи «генерация – эмоции – память - генерация». Далее в зависимости от характера потребности у индивида возникает желание, интерес или он борется с желанием, его психика подбирает наиболее удобные в данный период и при данном окружении «приличные» или рациональные мотивы и способ удовлетворения ощущаемой потребности.
Эта идея изложена или почти изложена в работе, посвященной годовщине П. К. Анохина [В. А. Правдивцев, С. Б. Козлов, Л. П. Нарезкина], в лекции студентам Смоленской государственной медицинской академии. В них комментируется этот же результат со ссылками на работы К. В. Судакова и Ю. А. Макаренко:
«По результатам исследований, проведенных К. В. Судаковым (1978, 1979), Ю. А. Макаренко (1980) было показано, что мотивационные возбуждения, находясь в цепи стабильной последовательности событий (потребность – мотивационные возбуждения – поведение, направленное на удовлетворение мотивации, – полезный результат) при многократном повторении этих событий начинают выступать сигналом будущего подкрепления. Следовательно, мотивационные возбуждения на самых ранних стадиях поведенческого акта уже закладывают основы формирования центрального комплекса возбуждений, предвосхищающего параметры будущего полезного поведенческого результата, т. е. акцептора результатов действия».
Все вышесказанное здесь просто подкрепляет с иной позиции уже изложенную почти как тридцать лет гипотезу К. В. Судакова и Ю. А. Макаренко.
Отсутствие сверхгенерации при активности. Если же сам субъект – носитель метапотребности – уже находится в той активности, которая и вызывает это положительное удовлетворение, то текущая генерация нейромедиаторов немедленно растрачивается и сбрасывается в процессе текущего эмоционального удовлетворения, при этом избытка медиаторов не возникает.
Итак, мы можем представить гипотезу активации метапотребности как процесс появления желания, потребности (метапотребности) с учетом всех моделей и гипотез на различных уровнях химико-биологическом, нейроинформационном и психологическом:
- на нейробиохимическом уровне – это экзоцитоз (излучение избытка эндогенно созревшего биологического материала опиоидного типа из секреторных оболочек и впрыскивание их в межсинаптическое пространство, что понижает пороговый уровень для прохождения импульса или вызывают генерацию случайного первичного импульса);
- на нейроинформационном уровне – это усиление возбуждения или связывания целых ансамблей-констелляций нейронов, сформированных на условно-рефлекторной основе и обеспечивающих формирование ощущаемой (индивидуально приобретенной) потребности;
- на психологическом уровне – это позитивное волнение, возбуждение, или положительное активирующее воспоминание, или спонтанное предвосхищение удовольствия (антиципация), которое постепенно приобретает черты осознанного мотива или желания и с нарастанием этого желания вплоть до формирования значимого пускового импульса (пусковой афферентации).
Все исследования формирования наркотического поведения в результате наркопотребности также применимы в данной системе. Отличие в следующем – метапотребность сформирована собственной биохимией организма и без подкрепления условного рефлекса внешними опиоидами. Вероятно, и это гипотеза автора, обычный человек не приносит себе большого вреда, получая эндогенное удовольствие от определенного вида деятельности потому, что собственные физиологические средства организма не могут привести к существенному нарушению биологических пределов. При этом мы знаем и отклонения от нормы, переходящие в патологию – навязчивые идеи, истощающее творчество и т.п. Иное дело в химической внешней наркомании. Условность рефлекса в наркомании состоит в том, что при употреблении определенного внешнего, т.е. экзогенного вещества человек получит удовольствие. Сама потребность формируется как смесь воздействия внешних опиоидов (их перманентного недостатка у наркомана) с внутренними опиоидами. Но внешние воздействия намного сильнее внутренних процессов автогенерации, и последние, какими бы не были, не могут восполнить отсутствие сильных внешних средств – возникает потребность во внешних уже испробованных источниках – зависимость становится не внутренней, а внешней. Но это не наша тема.
В этой связи мы можем и оппонировать взглядам авторов, отвергающим опиатную зависимость естественной эндогенной природы.
В связи с этим возникает интересный вопрос: зависимы ли мы от своих собственных энкефалинов и эндорфинов? Ответ на этот вопрос отрицательный… После высвобождения они достаточно быстро инактивируются, так что опиатные рецепторы оказываются занятыми только временно, и в результате толерантность и зависимость не развиваются. [Костерлиц Г.В. и Хьюс Д., 1981, с. 53-54].
Наша оценка такова. В приведенном мнении не учтена именно длительность процесса, в нем предполагается единократное воздействие. В реальности параллельно с инактивацией происходит непрерывная генерация или регенерация опиоида и новое высвобождение. Поэтому мы имеем постоянное раздражение рецепторов нейронов-получателей, их эндогенную настройку на генерацию везикул, на получение опиоидного раздражения. На психологическом уровне мы можем наблюдать в таких случаях и явное вербальное раздражение носителей такой зависимости («мне мешают работать!», «не мешай смотреть футбол!») и даже психозы. Опиатная зависимость возникает, и она становится потребностью. Совсем другое дела – она, как правило, не наносит ущерба здоровью индивидуума в отличие от внешней наркотической зависимости. Чаще, чисто внешне, метапотребность позволяет человеку выглядеть более счастливым в сравнении с людьми, озабоченными своими депривациями в материальном и статусном плане, что отмечал и Абрахам Маслоу. «Счастливые часов не наблюдают!»
Примеры запуска по аналогии с участием других потребностей (сексуальной потребности) – самопроизвольный оргазм во сне с включением эротических сновидений, как у мужчин, так и реже эротические сновидения у женщин.
Сны, как ассоциативное спонтанное возбуждение, галлюцинации в местах одиночного заключения (неудовлетворенная потребность общения) – все это примеры того же механизма, построенные на самых различных потребностях – безопасности (фобии, страхи – дурные сновидения), потребности общения.
В свете гипотезы о самопроизвольном излучении нейромедиаторов как метапотребности мы можем иначе посмотреть на некоторые известные факты, предубеждения и даже мифы.
Старые факты, идеи и даже предрассудки в новом свете
Давайте, вспомним, замечание Б.Ф. Поршнева в его «О начале человеческой истории», Глава 5, раздел «Палеолит и имитативный рефлекс» по поводу излишней работы троглодитида – австралопитека и архантропа – наш предок изготавливает сотни одинаковых рубил в количестве ему не нужном. И это еще, считает Поршнев, не человек, и это еще не труд, предполагая в этом просто импринтинг. В рамках выдвинутой здесь гипотезы мы можем дать некоторое иное объяснение палеоархеологическим данным. Здесь и обучение подроста этому занятию и удовольствие от большого количества «полезных» и просто «взрослых» вещей, имитация действий взрослой особи и, главное, мы можем предполагать удовольствие от этого «труда», как от творчества – изменения формы в первых своих видах труда, когда количество труда явно превышает потребность в его результатах. Запас рубил - это, возможно, и запас обороноспособности, степени защиты: безопасности 1 или 2. Позже мы снимем это предположение – человек в этот период (присваивающее хозяйство – охота) еще не умеет думать о будущем. Б. Ф. Поршнев критически относится к такой «бесполезной» деятельности, указывая на автоматизм. Думаю, он мыслит здесь как просто современный человек, уставший от рутины. Не стоит ли предположить, что преантроп уставал от многообразия и опасных неожиданностей, а не от рутины. Автоматизм тогда мог восприниматься как высшее интуитивное проявление красоты, порядка, удивительной предсказуемости в страшном и непредсказуемом мире. К этому же стоит отнести и процесс поддержания огня, почти единственный, которым овладел человек в природе, уже сам этот факт и достижение должны были вызывать бурный взрыв удовольствия, не менее, чем возню малыша со сборкой и разборкой пирамидки, и, скорее всего, именно на этом уровне сознания. Где уж нам осуждать автоматизм. Мы, в свою очередь, указываем, на роль эмоций, которые в добровольном подражании, несомненно, служат закреплению инструментального поведения. В чем можно быть точно уверенным, это в том, что наш праотец не заставлял, не принуждал прасына выделывать рубила.
Еще аргументы. Запущен не только импринтинг, имитативность, подражательность, о которой говорил Поршнев, но и игра в изготовление новых предметов – такая игра часто предполагает избыток сил и пищи у подроста, которого еще кормят старшие. Эта игра, как и импринтинг, свойственны, и раннему детству человека, и юному подросту у развитых биологических видов. Об интеллекте речи нет, но есть усвоение форм активности, и это усвоение не могло идти без эмоций. Изготавливаемые предметы – это инструменты добывания самой вкусной пищи для антропоида. И в России, в деревне, были традиции «бить баклуши» - заготовки для вырезания ложек – самая легкая работа для подростков, подозреваемых общиной в «идиотизме». Надо ли говорить, что подготовка рубил миллионы лет назад в плиоцене, задача не менее того. Рубило – важнейший инструмент для добычи мяса из трупа животного, особенно из мозговой коробки, куда не может добраться хищник, загнавший добычу и как хозяин, обглодавший ее. Только потом подходит к трупу наш слабовооруженный предок и начинает обрабатывать оставшееся. Так, что подготовка рубил не менее, чем изготовление ложки при воспоминании о еде, если предположить, что ребенку разрешат есть только ложкой, когда долгожданная еда появится. Какую роль может играть здесь предвосхищение употребления пищи? У сытого, возможно, игра? Еще не высокое творчество, а удовольствие хотя бы от движения. Дети часто повторяют одно и то же слово или делают одно и тоже действие. Для них это не рутинный, не скучный труд – это удовольствие от первых движений, все более точных движений. А у голодного человека? В воспоминании о будущей добыче? Вероятно, и рубила здесь играют самую прямую роль – именно, первую. У младенца или малыша, будь то человек или зверь, и у преантропа тоже – вся система потребностей как в зародыше заключена в один момент в одном движении, достаточно, например, посмотреть, как играет малышка в «дочки-матери», начиная с кормления, и кончая «школой».
Наконец, мы можем выйти на понимание и складывающегося инструментального разделения труда в сообществе людей. Человек с большим интересом делает то, что у него хорошо получается. Почему? Вовсе не для того, чтобы заработать, а именно потому, что хорошо получается! Это вызывает положительные собственные эмоции и, возможно, как дополнительное подкрепление, создатель получает похвалу от сожителей, соседей, членов, семьи или общины (охотников, земледельцев и т.п.). Все остальные механизмы вторичны и складываются позже. Поэтому нам следует относиться к положительной эмоции и к ее химико-физическому основанию, как к начальной (и конечной) обязательной фазе филогенетического развития человека, как к биологическому мостику между низшими потребностями и последующей инструментальной деятельностью человека по их удовлетворению.
Метапотребность как потребность, конкурирующая с обычными потребностями
Память о приятных воспоминаниях может улучшить состояние человека, оно же его может и расслабить (если это память об отдыхе). Но возникновение в памяти чего-то важного для человека, что может пригодиться в будущем, иногда поднимает (заставляет подниматься) засыпающего человека с постели – записать сейчас, чтобы не забыть утром.
Это означает, что мы в личном опыте имеем примеры того, как запускается важная для человека деятельность и становится пусковой афферентацией (исключительно на основе идей, которые приходят в голову). Если предположить, что нечто человек считает для себя постоянно важным (в психологии это может называться «интересом» и т.п.), то мысленная или мысленно воспринимаемая потребность может двигать человека очень существенно и даже в моменты, предназначенные для удовлетворения других потребностей (сон, голод и т.п.). Поэтому, сильная мотивация (не потребность) и рефлексия уже может, как пример, заместить на некоторое время другие потребности, например, более низкого уровня. Но тогда мы можем считать совершенно нормальным некоторый перекос баланса от низших, естественных потребностей в сторону метапотребностей – от деприваций - к удовольствиям или творчеству. Вспомним, что образ исследователя или ученого – это нечто «не от мира сего». Мы испытываем уважение к такому человеку. Он пренебрегает бытовыми проблемами, не отвлекаясь на них. Но его погруженность в абстрактную для общества тему не только «нравственный подвиг ради науки», как казалось нам, простым смертным. Природа сама, как бы, простите за поэтику, «поддерживает» такой альтруизм, «награждает» человека внутренней радостью, стимулирует его в поисках истины (правда, за его же энергетический счет). Сказанное никак не дает оснований пренебрегать и не ценить такое состояние и даже достояние, редкое пока в природе человечества. Его следует сохранять. О его преумножении - воспитании творчества в новых поколениях, о размерах потребного «духа» в конкретном обществе мы будем говорить отдельно.
Нашу нравственность, правда, может оскорблять то, что удовольствия (гедонизм в метапотребностях) могут приобретать простой, даже низменный характер. Но в этом мы ощущаем только общность закономерности (что народно, то не может не быть социальным фактом).
Но исключения есть не закон, а выброс, отклонение от закона как среднего, поскольку уже в биологии, в психологии человека и в социологии законы носят исключительно статистический характер. Итак, как правило, отклонения от нормы в системе потребностей – это, кроме собственно патологий, результат действия специфических, индивидуальных метапотребностей или просто сильных моментных эмоций.
Метапотребность как биологические часы
Особенно хорошо механизм метапотребностей наблюдается у людей, принявших «производственный» образ жизни, даже когда это не метапотребность, а просто установка на действие в хозяйственном цикле. Это автогенерация связей и даже напряженности организма по часам суток. Человек всю жизнь встает рано, чтобы идти на работу. Выйдя на пенсию, он сначала и довольно долго, а иногда и всю оставшуюся жизнь, может просыпаться рано и не может спать по утрам. Приобретенные биологические часы бодрствования и труда в процессе своего функционирования (постоянной работы) должны быть близки по типу к функционированию метапотребности: «организм зовет!». Он настроен на работу. Мы не можем иначе объяснить этим механизм как автоматическую генерацию определенных веществ в ЦНС индивида с учетом настройки его биологических часов. Запуск «готовности» к работе совершенно идентичен (и по нейрофизиологии) очередному запуску нового цикла метапотребности.
Метапотребность как плацебо
Обратимся к другим примерам. В шутливом определении менеджера у Т. Паркинсона позиция номер 1 представляет собою требование: менеджер – это тот, кто может заболеть в конце пятницы, но обязательно здоров к утру понедельника. Мы можем предполагать существование недельных или периодических часов, которые управляют мобилизацией иммунной и кратковременной мобилизацией защитных сил человека. Аналогичное можно установить по воспоминаниям актеров перед выступлением на сцене, преподавателей перед лекциями и т.п. К этому же разряду относят и плацебо, о котором довольно много сказано. Однако, нас интересует не единократное излучение адреналина и других нейромедиаторов и гормонов, а постоянное, или эпизодическое биохимическое действие такого рода.
До настоящего времени в литературе отмечалось значение нейромедиаторов и опиодов в качестве единократного плацебо. Например, указывался опыт африканских шаманов, которые лечили пациентов следующим образом. Его вводили в хижину, и в этот момент на него набрасывался средних размеров крокодил. Крокодила шаман оттаскивал, а многие пациенты после стресса чувствовали себя много лучше – полученный эмоциональный удар мобилизовал все силы, которые далее тратились на излечение основного заболевания. Но человек может использовать собственную метапотребность или направленность для ухода от физиологических трудностей или бытовых и жизненных лишений. Здесь не только «с милым рай и в шалаше», автор сам неоднократно проверял на себе – мысленное отвлечение на интересующие или творческие проблемы могут сделать течение времени незаметным, отключить на достаточно длительное время ощущение не только голода, но и холода (вне помещения). Кроме того, уже известно, что сильная положительная эмоция укрепляет иммунную систему.
Риск гибели от метапотребности при отсутствии возможности
удовлетворения
К классу опасных проявлений, построенных на регулярной самогенерации организмом биохимических средств борьбы или «химических средств защиты», следует отнести, например, нередкую гибель опытных и редко отдыхающих менеджеров в первый день – два своего отпуска. Организм, настроенный на работу, но никем не «предупрежденный», начинает обычный трудовой день, а человек валяется в постели (решил в кое-то веки выспаться!). Излучение нейрохимии должно быть встречено адекватной и хотя бы фальшивой нагрузкой или заторможено в первые дни химически.
Аутотренинги и самовнушение
В свете гипотезы следует вывод, что аутотренинг и самовнушение – это принудительная перестройка нейронной сети. Если мы хотим, чтобы она закрепилась всерьез и надолго, ее следует подкреплять возможно более сильным положительным эмоциональным состоянием. В этой связи народный опыт дает интересные решения – баня, парная, сауна – это закалка и в целом нагрузка. Но если она сопровождается встречей с друзьями и даже умеренным возлиянием, то становится не просто традицией, а полезной потребностью, причем друзья и хорошие эмоции, как показывает хотя бы телевизионный опыт фильма «С легким паром», много важнее.
Метастрахи и метазапреты– фобии и неврозы
Неврозы и различные запреты возникают исключительно как отрицательный личный или наблюденный чужой опыт, растиражированный воображением и закрепленный в нейросистеме индивида. Понимание пациентом социальной нормы и чисто техническое рациональное отношение к «кривой», т.е. неверной, настройке собственной нервной системы способно было бы исправить множество патологий, успокоить и полноценно «отремонтировать» психику миллионов людей. Это делается малыми шагами и непрерывными положительными подкреплениями – в ход следует пускать самые большие доступные радости.
Молекулы «радости», «страха» или «агрессии»
Ряд современных исследователей, в частности, в современной России, например, В.Шестаков, руководитель Научно-учебного центра проблем жизнедеятельности человека РАН [Шестаков В., 2003], поставили целью вводить животных в определенные состояния, брать их нейропептидные компоненты из крови и сыворотки и вводить в организм пациентов в лечебных и психологических корректирующих целях. Известно, впрочем, что состав таких вытяжек из нейромедиаторов и нейрогормонов весьма сложен. Небольшое изменение концентрации компонент приводят к совершенно другим результатам или мало эффективно.
В этой связи очень интересно вспомнить историко-религиозную или скорее культурную традицию «кошерности» у евреев как отражение их длительного опыта скотоводов, до оседания в Палестине. Поедание мяса испуганного и долго убиваемого зверя (домашнего или на охоте) означает физически включение в пищу и нейромедиаторов в комбинации страха и стресса (особенно в свежей крови, которую в пустыне или в безводной степи скотоводы могли пить свежей из артерии вместо воды). Пастыри, вожди колен Израилевых или первосвященники, вероятно, заметили, какие-то отрицательные воздействия от употребления в пищу такого мяса на свою паству и наложили запрет. Это никак не противоречит различным другим народным традициям и древним мифам, например, рекомендациям убить в открытом бою хищного зверя и съесть его сердце, что может «придать храбрости» и т.п., наоборот, согласуется с ними.
Изложенная (на май 2004 г.) гипотеза активации метапотребности имеет целью поддержать теорию иерархии потребностей Маслоу как общую теорию потребностей и мотивации. Мы с удовольствием укажем на материалы других исследователей, работающих или получивших результаты в этом направлении
Выводы раздела 4
Даны современные представления о физико-химическом механизме формирования нейронных связей как отражении вышеописанных психологических процессов настройки и запоминания положительных эмоций и формирования в последующем метапотребностей. На этой основе выдвинута и обосновывается гипотеза о формировании метапотребностей с помощью эмоциональных положительных подкреплений и их спонтанное проявление через самопроизвольный экзоцитоз опиоидных нейромедиаторов.
Литература к разделу 4
Анохин П.К., Системный анализ ингегративной деятельности нейрона, под. ред, П.К, Анохина, М., Наука, 1974.- 159 с.
Анохин П.К. Системные механизмы высшей нервной деятельности: избр. тр. - М.: Наука, 1979. – 453 с.
Ашмарин И.П., Бородкин Ю.С. и др., Механизмы памяти, рук-во по физиологии, отв. ред. Г.А. Вартанян, Л., Наука – ЛО, 1987, - 432 с.
Гомазков О.А., Нейропептиды – универсальные регуляторы. Почему?, Природа, 1999, № 4. http://www.ibmh.msk.su/vivovoco/VIVOVOCO.HTM
Зефиров А.Л., 2000, Биология
Колесов Д.В. Введение в общую психологию, уч. пос., Воронеж, 2002.
Кольман Я., Рем К.-Г., Наглядная биохимия. http://biochemistry.vov.ru/nagl_bio/index.htm
Костерлиц Г.В. и Хьюс Д., Развитие концепций опиантных рецепторов и их лигандов, с.43-55. в сб. ст. Эндорфины, под ред. Э.Коста и М. Трабукки, пер. с англ. К.б.н. М.А. Панова, М., «Мир», 1981.- 386 с.
Макаренко Ю. А. Системная организация эмоционального поведения. - М.: Медицина,1980. - 157 с.
Немов Р.С. Психология, кн. 1. Общие основы психология. – 2-е изд. – М.: Просвещение: ВЛАДОС, 1995. – 576 с.
Поршнев Б.Ф. О начале человеческой истории. Проблемы палеопсихологии.
– М., 1974.
Правдивцев В.А., Козлов С.Б., Нарезкина Л. П., Архитектура поведенческого акта с позиции теории функциональной системы П.К. Анохина, лекции, Смоленская государственная медицинская академия.1998.
Стайн Л., Беллуци Дж., Эндорфины мозга и ощущение хорошего самочувствия: психологическая гипотеза, с.294-307 в сб. ст. Эндорфины, под ред. Э.Коста и М. Трабукки, пер. с англ. К.б.н. М.А. Панова, М., «Мир», 1981.- 386 с.
Судаков К. В. Биологические мотивации. - М.: Медицина,1971. - 303 с.
Судаков К. В. Общая теория функциональных систем. - М.: Медицина, 1984.-224 с.
Судаков К. В. Функциональные системы организма. Под ред. К. В. Судакова. -М.: Медицина, 1987.- 432 с.
Судаков К. В. Избранные лекции по нормальной физиологии.-М.:Эрус,1992.-243 с.
Трифонов Е.В. Психофизиология человека, толковый русско-английский словарь, 2001.
Шестаков В., Шекспир в пробирке, Огонек, Февраль 2003, № 5 (4784).
Эндорфины, под ред. Э.Коста и М. Трабукки, сб. ст., пер. с англ. к.б.н. М.А. Панова, М., «Мир», 1981.- 386 с
Автор использовал информацию сайтов:
http://www.it-med.ru/index.php
http://max.1gb.ru/main/main.shtml
http://www.bio.msu.ru/physiology225/index.htm
http://biochemistry.vov.ru/nagl_bio/index.htm
http://courier.com.ru/priroda/pricont.htm
http://www.manualtherapy.agava.ru/
http://ndolya.boom.ru/prometheus/kp17.htm
http://www.glossary.ru/index.htm
http://tryphonov.narod.ru/tryphonov/auth_r.htm
http://www.biohome.ru/news.php
http://globus.smolensk.ru/user/sgma/MMORPH/N-3-html/3.htm

{kind=link}